Language
Login
Language Setting
X
English
日本語 [Japanese]
about this App
Non-official example of GenoCon
useful
5
Loading...
//--------------------------------------------------------------- // window.onloadメソッド //概要: // 遺伝情報の組み立ての主処理。 // 本関数内で使用している独自に定義したオブジェクトDNAとProteinは // 本コード後半で定義している。 //引数: // なし //戻り値: // なし //--------------------------------------------------------------- window.onload = function() { $( '#result' ).append( '<h1>GenoCon Example</h1>' ); $( '#result' ).append( '<p class="message">Start!</p>' ); try { //HPSタンパク質のアミノ酸配列の読み込み var HPS = new Protein(); HPS.loadSequence( 'rdf1s222i', 'protein', 'http://www.uniprot.org/uniprot/Q9LBW4' ); $( '#result' ).append( '<h2>HPS:</h2>'); HPS.write(); //PHIタンパク質のアミノ酸配列の読み込み var PHI = new Protein(); PHI.loadSequence( 'rdf1s222i', 'protein', 'http://www.uniprot.org/uniprot/Q9LBW5' ); $( '#result' ).append( '<h2>PHI:</h2>' ); PHI.write(); //HPSとPHIの融合タンパク質の生成 var HPSPHI = new Protein( HPS, PHI ); $( '#result' ).append( '<h2>HPSPHI:</h2>'); HPSPHI.write(); //プラスチド移行シグナルペプチドの付加 HPSPHI.addSignalPeptide( 'http://scinets.org/item/cria228s2ria228s4i' ); $( '#result' ).append( '<h2>signal+HPSPHI:</h2>' ); HPSPHI.write(); //逆翻訳 var hpsphi = HPSPHI.reverseTranslate( 'rdf1s291i', 'codon_table_of_Arabidopsis_thaliana' ); $( '#result' ).append( '<h2>reverseTranslated signal+HPSPHI (hpsphi gene):</h2>' ); hpsphi.write(); //IRESの読み込み var ires = new DNA(); ires.loadSequence( 'rdf1s222i', 'dna', 'http://scinets.org/item/cria228s1ria228s1i' ); $( '#result' ).append( '<h2>IRES:</h2>' ); ires.write(); //添付ファイルからのepfl9遺伝子の塩基配列の読み込み var epfl9 = new DNA(); epfl9.loadSequence( 'rdf1s286i', 'epfl9_gene', 'http://www.ebi.ac.uk/ena/data/view/BAD43639' ); $( '#result' ).append( '<h2>epfl9:</h2>' ); epfl9.write(); //融合タンパク質の遺伝子、IRES、epfl9の塩基配列の結合 var finalSeq = new DNA( hpsphi, ires, epfl9 ); $( '#result' ).append( '<h2>hpsphi+IRES+epfl9:</h2>' ); finalSeq.write(); $( '#result' ).append( '<p>length = ' + finalSeq.getLength() + ' bp</p>'); } catch( e ) { $( '#result' ).append( '<p class="error">' + e + '</p>' ); } finally { $( '#result' ).append( '<p class="message">successfully-completed!<p>' ); } }; //////////////////////////////////////////////////////////////////////////////////// // // 以下は遺伝情報の組み立てのための汎用的モジュール // //////////////////////////////////////////////////////////////////////////////////// //--------------------------------------------------------------- //Sequenceクラスのコンストラクタ //概要: // DNA、Proteinなどのシーケンスを格納・管理するための // クラスの親クラス。Sequenceクラス自体のインスタンスは生成せず // 抽象クラスとして扱うものとする。 //引数: // なし //戻り値: // なし //--------------------------------------------------------------- function Sequence() { //Sequenceクラスは抽象クラス } //--------------------------------------------------------------- //Sequenceクラスのメンバ _checkSequenceメソッド //概要: // シーケンスに既定のアルファベット(塩基やアミノ酸)以外の文字 // が含まれていないか検査し、その場合はエラーを出す。 //引数: // seq 検査のシーケンス対象となるシーケンスの文字列 //戻り値: // なし //--------------------------------------------------------------- Sequence.prototype._checkSequence = function( seq ) { var i; for( i = 0; i < seq.length; i++ ) { var j; for( j = 0; j < this.constructor._alphabet.length; j++ ) { if( seq.charAt( i ) === this.constructor._alphabet[ j ] ) { break; } } if( j === this.constructor._alphabet.length ) { throw new Error( 'シーケンスデータに不正なアルファベット' + seq.charAt( i ) + 'が含まれています。' ); } } }; //--------------------------------------------------------------- //Sequenceクラスのメンバ setSequenceメソッド //概要: // オブジェクトにDNAやタンパク質のシーケンスをセットする。 // 複数の引数を与えるとそれらの配列を準に連結してセットする。 //引数: // arg シーケンスの文字列か、同じクラスのオブジェクト //戻り値: // なし //--------------------------------------------------------------- Sequence.prototype.setSequence = function( arg ) { var sum = ''; var i; for( i = 0; i < arg.length; i++ ) { if( this.constructor !== arg[ i ].constructor && ( typeof arg[ i ] ) !== 'string' ) { throw new Error( '文字列以外の値あるいは異なるクラスのオブジェクトが引数として与えられました。' ); } sum += arg[ i ].toString().replace( /\s/g, '' ); } this._checkSequence( sum ); this._sequence = sum; }; //--------------------------------------------------------------- //Sequenceクラスのメンバ loadSequenceメソッド //概要: // シーケンスをLinkDataのテーブルデータから取得して、オブジェクトにセットする。 // 基本的にはLinkData.getObjectsのwrapperだが、 // propertyの#以前の省略に対応。(例: http://linkdata.org/property/rdf1s222i#Sequence → #Sequence) // さらにpropertyが省略された場合には、http://linkdata.org/property/[WORK ID]#Sequenceをpropertyとする。 //引数: // workID LinkDataの作品のID // filename 作品中のデーブルのファイル名 // subject 主語 (表の各行の見出しに相当) // property 術後 (表の各列の見出しに相当) //戻り値: // なし //--------------------------------------------------------------- Sequence.prototype.loadSequence = function( workId, filename, subject, property ) { var seq; var emp = []; if( property === undefined ) { property = '#Sequence'; } if( property.charAt( 0 ) === '#' ) { property = 'http://linkdata.org/property/' + workId + property; } seq = LinkData.getObjects( workId, filename, subject, property ); if( seq[0] === undefined ) { throw new Error( 'LinkDataからの配列の取得に失敗しました。' ); } this.setSequence( seq ); }; //--------------------------------------------------------------- //Sequenceクラスのメンバ getSequenceメソッド //概要: // オブジェクトにセットされたシーケンスを取得する //引数: // なし //戻り値: // シーケンスの文字列 //--------------------------------------------------------------- Sequence.prototype.getSequence = function() { if( this._sequence === undefined ) { throw new Error( 'シーケンスがセットされていないオブジェクトのシーケンスが参照されました。' ); } return this._sequence; }; //--------------------------------------------------------------- //Sequenceクラスのメンバ toStringメソッド //概要: // シーケンスを文字列で返す。getSequenceを流用しているだけだが、 // これによってオブジェクトを文字列的に扱える。 // var P = new Protein( 'ACDEFG' ); // alert( P ) // なし //引数: // なし //戻り値: // シーケンスの文字列 //--------------------------------------------------------------- Sequence.prototype.toString = function() { return this.getSequence(); }; //--------------------------------------------------------------- //Sequenceクラスのメンバ writeメソッド //概要: // シーケンスを画面に出力。 //戻り値: // なし //--------------------------------------------------------------- Sequence.prototype.write = function() { var output = ''; var i; var seq = this.getSequence(); $( '#result' ).append( '<div class="sequence">' + seq + '</div>' ); }; //--------------------------------------------------------------- //Sequenceクラスのメンバ getLengthメソッド //概要: // シーケンスの長さを返す。 //引数: // なし //戻り値: // シーケンスの長さ //--------------------------------------------------------------- Sequence.prototype.getLength = function() { return this.getSequence().length; }; //--------------------------------------------------------------- //Sequenceクラスの子クラス DNAクラスのコンストラクタ //概要: // DNAのシーケンス(塩基配列)を格納・管理するためのクラス //引数: // 元になるDNAオブジェクトあるいはシーケンスの文字列(複数指定可) //戻り値: // なし //--------------------------------------------------------------- function DNA() { this.constructor = DNA; if( arguments.length > 0 ) { this.setSequence( arguments ); } } // Sequenceを継承 DNA.prototype = new Sequence(); //各ヌクレオチドをDNAを構成するアルファベットとして設定。 DNA._alphabet = [ 'A', 'T', 'G', 'C' ]; //--------------------------------------------------------------- //Sequenceクラスの子クラス Proteinクラスのコンストラクタ //概要: // タンパク質のシーケンス(アミノ酸配列)を格納・管理するためのクラス //引数: // 元になるProteinオブジェクトあるいはシーケンスの文字列(複数指定可) //戻り値: // なし //--------------------------------------------------------------- function Protein() { this.constructor = Protein; if( arguments.length > 0 ) { this.setSequence( arguments ); } } // Sequenceを継承 Protein.prototype = new Sequence(); //アミノ酸と終止コドンに対応する*をタンパク質を構成するアルファベットとして設定。 Protein._alphabet = [ 'A', 'C', 'D', 'E', 'F', 'G', 'H', 'I', 'K', 'L', 'M', 'N', 'P', 'Q', 'R', 'S', 'T', 'Y', 'V', 'W', '*' ]; //--------------------------------------------------------------- //Proteinクラスのメンバ addSignalPeptideメソッド //概要: // シーケンスにシグナルペプチドを不可する。 //引数: // subject signal peptide (http://linkdata.org/work/rdf1s292i/) // のテーブルデータsignal_peptideのシグナルペプチドを示す主語。 // position 挿入位置不定の場合はシグナルペプチドの挿入位置。 // (何番目のアミノ酸残基の後に挿入するか) // シグナルペプチドのシーケンス中に不特定部分XXXを含む場合には、 // 場合は3番目以降)の引数で順にXXX部分に入るシーケンスを指定する。 // 例) // N末端挿入型シグナル XXXXRLXXXXXHL-の始めのXXXXをAAAA // 次のXXXXXをPPPPPとして付加する場合: // HPSPHI.addSignalPeptide( 'http://scinets.org/item/cria228s2ria228s8i', 'AAAA', 'PPPPP' ); // 不特定部位挿入型シグナル -PPKKKRKV-を10残基目のアミノ酸残基の // 直後に挿入する場合: // HPSHPI.addSignalPeptide( 'http://scinets.org/item/cria228s2ria228s1i', 10 ); //戻り値: // なし //--------------------------------------------------------------- Protein.prototype.addSignalPeptide = function( subject, position ) { //指定されたシグナルペプチドの読み込み var signalPeptide = LinkData.getObjects( 'rdf1s292i', 'signal_peptide', subject, 'http://linkdata.org/property/rdf1s292i#sequence' ).toString(); //挿入部位の情報を読み込み var location = LinkData.getObjects( 'rdf1s292i', 'signal_peptide', subject, 'http://linkdata.org/property/rdf1s292i#location' ).toString(); //シグナルペプチド中のXXX部分の置き換えに用いる引数の開始位置。 //末端に挿入されるシグナルペプチドなら2番目以降の引数が用いられるが、 //不特定の場合には2番目の引数は挿入位置を示すため、3番目以降になる。 var start = 2; var inputSeq = this.getSequence(); var outputSeq; var i; //シグナルペプチドから挿入部位を示すハイフンを除去 signalPeptide = signalPeptide.replace( /-/g , '' ); switch( location ) { //N末端 // 元データの綴り間違いと正しい綴りの両方に対応 case 'N-terminus': case 'N-teminus': outputSeq = signalPeptide+inputSeq; break; //C末端 case 'C-terminus': outputSeq = inputSeq+signalPeptide; break; //不特定 case 'Any': if( position > inputSeq.length || position < 0 ) { throw new Error( 'シグナルペプチドの挿入位置が不正です。(挿入位置:' + position + ')' ); } outputSeq = inputSeq.slice( 0, position ) + signalPeptide + inputSeq.slice( position, inputSeq.length ); start = 3; break; } //シグナルペプチドに未確定のXを含む場合には、その部分に2番目以降 //(挿入位置が不特定の場合には3番目)の引数を順番に当てはめていく。 for( i = start - 1; i < arguments.length; i++ ) { outputSeq = outputSeq.replace( /X+/i , arguments[ i ] ); } this._checkSequence( outputSeq ); this._sequence = outputSeq; }; //--------------------------------------------------------------- //Proteinクラスのメンバ _getGeneticCodeTableメソッド //概要: // コドン頻度表を取得し、連想配列に格納。 //引数: // workId LinkData上のコドン頻度表のwork ID // filename LinkData上のコドン表のファイル名 //戻り値: // コドン頻度表を格納した連想配列 //--------------------------------------------------------------- Protein.prototype._getGeneticCodeTable = function( workId, filename ) { //遺伝暗号表を格納する連想配列 var geneticCodeTable = {}; //一時的にあるアミノ酸に対応するコドンを添字にしてその頻度を格納する連想配列 var fraction = {}; var subjects = LinkData.getSubjects( workId, filename ); var i; for( i = 0; i < Protein._alphabet.length; i++ ) { var j; for( j = 0; j < subjects.length; j++ ) { var aa = LinkData.getObjects( workId, filename, subjects[ j ], 'http://linkdata.org/property/' + workId + '#one-letter%20code%20of%20amino%20acid' ).toString(); if( aa === Protein._alphabet[ i ] ) { var codon= subjects[ j ].toString().split( '#' )[1]; fraction[ codon ] = LinkData.getObjects( workId, filename ,subjects[ j ], 'http://linkdata.org/property/' + workId + '#usage' ).toString(); } } geneticCodeTable[ Protein._alphabet[ i ] ] = fraction; fraction = {}; } return geneticCodeTable; }; //--------------------------------------------------------------- //Proteinクラスのメンバ _DHondtメソッド //概要: // 比例代表選挙の各政党への議席数の割り振りに用いられる // ドント方式を応用して、与えられたコドンの使用頻度から // タンパク質の逆翻訳時に使用される各コドンの個数を算出し、 // その情報を配列に格納して返す。 //引数: // totalSeatsNum タンパク質を構成するあるアミノ酸の総数(総議席数に相当) // ratio 各コドンの頻度を含む連想配列(得票数に相当) //戻り値: // コドンを添え字にし、そのコドンの使用数を持つ連想配列 //--------------------------------------------------------------- Protein.prototype._DHondt = function( totalSeatsNum, ratio ) { var max; var maxParty; var gottenSeats = {}; var divisor = {}; var i; for( i in ratio ) { if( ratio.hasOwnProperty( i ) ) { gottenSeats[ i ] = 0; divisor[ i ] = 1; } } for( i = 0; i < totalSeatsNum; i++ ) { var party; max = 0; for( party in ratio) { if( ratio.hasOwnProperty( party ) ) { if( max < ( ratio[ party ] / divisor[ party ] ) ) { maxParty = party; max = ratio[ maxParty ] / divisor[ maxParty ]; } } } gottenSeats[ maxParty ]++; divisor[ maxParty ]++; } return gottenSeats; }; //--------------------------------------------------------------- //Proteinクラスのメンバ _getAmniAcidCompositionメソッド //概要: // シーケンス中に各アミノ酸が何残基ずつ存在するかを返す。 //引数: // inputSeq 対象になるシーケンスの文字列 //戻り値: // アミノ酸を添え字にし、そのアミノ酸がシーケンス中で // 使用された回数を持つ連想配列 //--------------------------------------------------------------- Protein.prototype._getAminoAcidComposition = function( inputSeq ) { var numOfAA = {}; var i; //配列AAnumの初期化 for( i = 0; i < Protein._alphabet.length; i++ ) { numOfAA[ Protein._alphabet[ i ] ] = 0; } for( i = 0; i < inputSeq.length; i++ ) { numOfAA[ inputSeq.charAt( i ) ]++; } return numOfAA; }; //--------------------------------------------------------------- //Proteinクラスのメンバ _getIdealCodonCompotitionメソッド //概要: // タンパク質のアミノ酸組成とコドンの使用頻度に基づいて、 // 各コドンの最適な使用回数を算出する。 //引数: // geneticCodeTable 頻度情報を含むコドン表のデータ // inputSeq 対象になるシーケンスの文字列 //戻り値: // アミノ酸を添え字にし、そのアミノ酸の最適な使用回数を持つ連想配列 //--------------------------------------------------------------- Protein.prototype._getIdealCodonCompotition = function( geneticCodeTable, inputSeq ) { var numOfAA = this._getAminoAcidComposition(inputSeq); var idealCompo = {}; var aa; for( aa in geneticCodeTable ) { if( geneticCodeTable.hasOwnProperty( aa ) ) { idealCompo[ aa ] = this._DHondt( numOfAA[ aa ], geneticCodeTable[ aa ] ); } } return idealCompo; }; //--------------------------------------------------------------- //Proteinクラスのメンバ _getIdealCodonCompotitionメソッド //概要: // タンパク質を逆翻訳し、結果をDNAオブジェクトで返す。 //引数: // workId LinkData上のコドン頻度表のwork ID // filename LinkData上のコドン表のファイル名 //戻り値: // 逆翻訳結果のシーケンスが格納されたDNAオブジェクト //--------------------------------------------------------------- Protein.prototype.reverseTranslate = function( workId, filename ) { //遺伝暗号表の取得 var geneticCodeTable = this._getGeneticCodeTable( workId, filename ); //終止コドン付加 var inputSeq = this.getSequence() + '*'; //最適なコドンの使用頻度の取得 var idealComposition = this._getIdealCodonCompotition( geneticCodeTable, inputSeq ); //あるアミノ酸に対応するコドンを一時的に格納する配列 var codons = []; //あるアミノ酸ごとに使用するコドンを使用する個数だけ使用する順に詰めたキュー配列 var tQueue = []; //アミノ酸ごとにtQueueの内容を格納したキュー配列 var codonQueue = {}; //コドンを使用頻度順にソートするための関数 var sortCodons = function( a, b ) { return b.number - a.number; }; //塩基配列を書き出す先 var outputSeq = ''; var aa; for( aa in idealComposition ) { if( idealComposition.hasOwnProperty( aa ) ) { var codon; var i; for( codon in idealComposition[ aa ] ) { if( idealComposition[ aa ].hasOwnProperty( codon ) ) { //コドンとその頻度を配列に格納 codons.push( { 'codon' : codon, 'number' : idealComposition[ aa ][ codon ] } ); } } //使用頻度順にコドンをソート codons.sort( sortCodons ); //tQueueに使用する順にコドンを詰め込む for( i = 0; i < codons.length; i++ ) { var j; for( j = 0; j < codons[ i ].number; j++ ) { tQueue.push( codons[ i ] . codon ); } } codonQueue[ aa ] = tQueue; codons = []; tQueue = []; } } //実際にコドンを並べていく。 for( i = 0; i < inputSeq.length; i++ ) { outputSeq += codonQueue[ inputSeq.charAt( i ) ][ 0 ]; codonQueue[ inputSeq.charAt( i ) ].shift(); } return new DNA( outputSeq ); };
.sequence { font-family: monospace;word-wrap: break-word; } .error { color: red; font-weight: bold; font-size:large } .message { color: green; font-weight: bold; font-size:large } h2 {border-bottom: 1px solid #333399}
<div id="result"></div>
Preview
Input Data
ReadMe
Snapshots
LinkData Work
Table Data
Codon usage table of Arabidopsis thaliana
Contributor:Masahiro Mochizuki
Update:Sep 27, 2012
860 Downloads, 3 Applications
アミノ酸は一文字表記で記した。なお、終止コドンはアミノ酸をコードしないため、コードするアミノ酸の一文字表記の代わりに*で示した。
codon_table_of_Arabidopsis_thaliana
GenoCon2 ChallengeB sample data
Contributor:GenoCon
Update:Sep 21, 2012
3354 Downloads, 8 Applications
dna
promoter
protein
signal_peptide
terminator
シグナルペプチド
Contributor:Masahiro Mochizuki
Update:Sep 27, 2012
690 Downloads, 3 Applications
シグナルペプチドの一覧。 プロパティ名は必ずしも直感的でないが、出典に従った。また、locationプロパティでN末端にーを意味するオブジェクトがN-teminusとなっているが原文ママ。
signal_peptide
Add LinkData work(LinkData)
Link http://app.linkdata.org/run/app1s63i?tab=readme
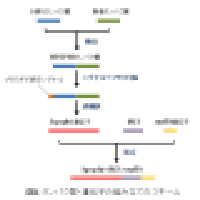
<p></p><div id="swf_body"> <div id="wiki"><br><h2>1. プログラムの流れ</h2><blockquote>(1) <em>Mycobacterium gastri</em>のヘキスロース6-リン酸合成酵素(<strong>HPS</strong>) のアミノ酸配列を取得。<br>(2) <em>Mycobacterium gastri</em>のヘキスロース6-リン酸イソメラーゼ(<strong>PHI</strong>) のアミノ酸配列を取得。<br> (3) <strong>HPS</strong>と<strong>PHI</strong>のアミノ酸配列を融合して融合タンパク質 <strong>HPSPHI</strong>を生成。<br>(4) <strong>HPSPHI</strong>にプラスチド移行シグナルを付加。<br>(5) シグナルペプチドが付加された<strong>HPSPHI</strong>を逆翻訳し、<strong>hpsphi</strong>遺伝子を生成。<br>(6)内部リボソーム進入部位 (IRES)の塩基配列を取得。<br>(7) シロイヌナズナのepidermal patterning factor-like 9 遺伝子(<strong>epfl9</strong>)の塩基配列を取得。<br>(8) <strong>hpsphi</strong>+<strong>IRES</strong>+<strong>epfl9</strong>の順で塩基配列を結合。<br><br></blockquote><blockquote><a><img src="https://database.riken.jp/sw/download/cria182s4ria182s97i%7Efiles%7Egenocon2010.png?&size=100" title="cria182s4ria182s97i~files~genocon2010.png" class="image "></a></blockquote><br><br><h2>2. デザインの狙い</h2><blockquote> 本デザインは、ヘキスロース6-リン酸合成酵素(HPS)とヘキスロース 6-リン酸イソメラーゼ(PHI)からなる融合タンパク質を発現させ、ホルムアルデヒドをカルビン回路に取り込ませる形で吸収させることを期待したもので ある。これは、先行研究[1,2]よって実証されている方法をヒントにしたものだが、以下に示す独自の工夫によって吸収効率の向上を目指し た。<br></blockquote><h4>2-1-1. 独自のコドン最適化アルゴリズム</h4><blockquote> 生物種によってゲノ ム全体でのコドンの使用頻度には偏りがあり、その偏りは主に各コドンに対応するtRNAの細胞内濃度を反映していることが知られている[3]。この性質の ため異種由来の遺伝子を宿主で高発現させるには、宿主細胞内の特定のtRNAが不足することがないように、宿主のコドンの使用頻度を考慮して遺伝子を構成 するコドンを同義コドンに置き換える処理(コドン最適化)を行うことが有効とされる[4]。<br>私が開発したアルゴリズムでは、まず高発現させたいタ ンパク質のアミノ酸組成を算出し、各アミノ酸についてどのコドンを何回使えば宿主であるシロイヌナズナのコドン使用頻度に最適合するかを算出する。次に、タンパク質のアミノ酸配列に従って、高頻度のコドンか ら順に5'から3'末端の方向にコドンを並べていくことで遺伝子の塩基配列を構築する。これは、5'末端側のコドン最適化が最も効果的だという報告[5] があるためで、この方法なら対応tRNAが豊富なコドンが5'末端側に優先的に割り当てられ、5'領域により重点的な最適化を実現できる。<br></blockquote><h4>2-1-2. 気孔の増加によるホルムアルデヒド吸収の効率化の試み</h4><blockquote> ホルムアルデヒドはシックハウス症候群の原因物質の一つであり、実用的な場面での吸収のターゲットを空気中のホルムアルデヒドだと想定した。また、今回採 用したホルムアルデヒドの吸収の方法は光合成反応の一部であるカルビン回路を利用したものであり、この回路に取り込まれるホルムアルデヒドの大部分は二酸 化炭素などと同じく、葉の表面の気孔を通じて主たる光合成反応の場である葉肉に運ばれると思われる。そこで、私は気孔の数を増やすことでホルムアルデヒド の植物体への取り込みを容易にし、ホルムアルデヒド吸収効率を向上させようと考えた。<br> そのために本デザインではepfl9遺伝子を過剰発現する方法を採用した。epfl9は<em>STOMAGEN</em>とも呼ばれ、シロイヌナズナで過剰発現させると気孔の数を増加させることが実験的に証明されている[6]。<br></blockquote><h2>5. 参考文献</h2><ol><li>L.-m. M. Chen, <em>et al</em>. , <em>Bioscience, biotechnology, and biochemistry</em> <strong>74</strong>, 627 (2010). <a href="http://view.ncbi.nlm.nih.gov/pubmed/20208346">[Pubmed]</a></li><li>I. Orita, N. Sakamoto, N. Kato, H. Yurimoto, Y. Sakai, <em>Applied microbiology and biotechnology</em> <strong>76</strong>, 439 (2007). <a href="http://view.ncbi.nlm.nih.gov/pubmed/17520247">[Pubmed]</a></li><li>T. Ikemura, <em>Mol Biol Evol</em> <strong>2</strong>, 13 (1985). <a href="http://view.ncbi.nlm.nih.gov/pubmed/3916708">[Pubmed]</a></li><li>C. Gustafsson, S. Govindarajan, J. Minshull, <em>Trends in biotechnology</em> <strong>22</strong>, 346 (2004). <a href="http://view.ncbi.nlm.nih.gov/pubmed/15245907">[Pubmed]</a></li><li>E. B. Vervoort, <em>et al</em>. , <em>Nucleic acids research</em> <strong>28</strong>, 2069 (2000). <a href="http://view.ncbi.nlm.nih.gov/pubmed/10773074">[Pubmed]</a></li><li>S. S. Sugano,<em> et al</em>. ,<em> Nature</em> <strong>463</strong>, 241 (2010). <a href="http://www.citeulike.org/user/Mochimasa/article/6430744">[Pubmed]</a></li></ol></div> </div> <p></p>
jquery-1.7.1.min.js
Work
Add
Clear
insert work id or work name.